INTRODUCTION
Advances in surgical and medical management of acutely injured individuals, such as decompression within 24 hours and hemodynamic management, have decreased mortality and improved neurologic outcomes from spinal cord injury (SCI) [1-5]. Additionally, strides in technologic and therapeutic development hold great promise for the future of SCI recovery [6-17]. Despite this, a cure for disabling sensorimotor deficits caused by SCI remains elusive for the nearly 18,000 individuals who experience SCI each year. While the most obvious effects of SCI are physical, the emotional and financial well-being of individuals and their families are also impacted. With rehospitalization rates as high as 36% [18], the first year costs of living with SCI range from $400,000 to more than $1.2 million depending on level and severity [19]. Importantly, gains in mobility reduce both rehospitalization18 and overall lifetime costs [20] and ultimately improve individuals’ quality of life [21]. As such, there is a critical unmet clinical need for acute interventions that can jumpstart recovery of functional mobility.
Activity-based rehabilitation paradigms use principles of motor learning and neuroplasticity, such as repetition, specificity, and intensity in an effort to minimize compensation and promote recovery of function after neurologic insult [22-25]. To date, these rehabilitation approaches have demonstrated significant promise for promoting functional recovery after SCI and will likely be vital adjuncts to pharmacological, cellular, or technological interventions in the future [6-17,26,27]. However, despite the fact that rehabilitation may be required to guide neuroplasticity and harness the beneficial effects of novel therapeutics, standardized physical therapy regimens are rarely included when studying pharmacological interventions [6,28-32]. Moreover, despite intensive rehabilitation, recovery from SCI is often incomplete [33-35]. As such, additional understanding of the neurobiology of SCI and recovery, both natural and training-induced, is essential to maximize outcomes.
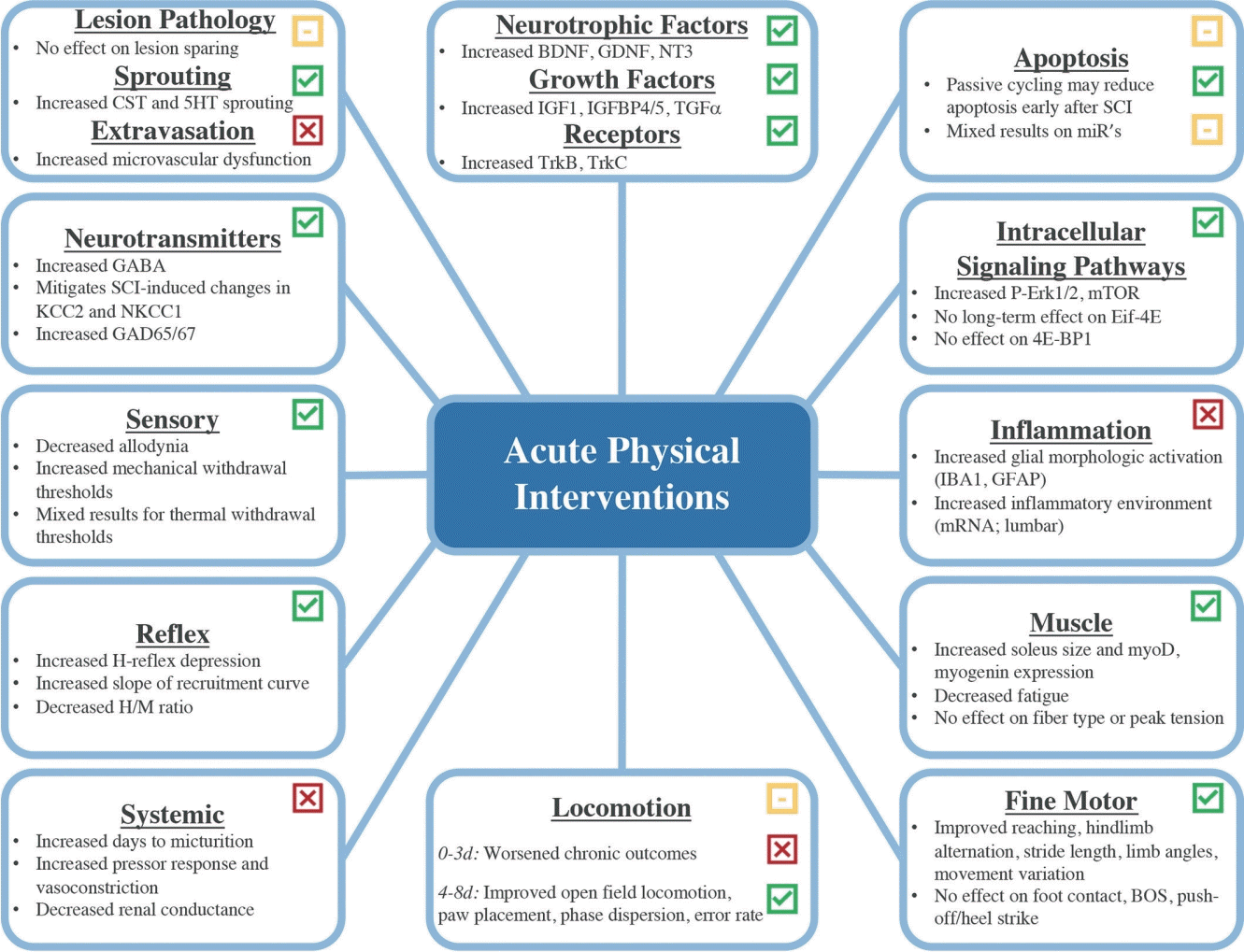
The timeliness of physical intervention may improve functional outcomes after SCI, but our understanding of this effect is incomplete, and results are mixed. Indeed, while some studies indicate a positive effect of early physical intervention [36-38] others suggest that early intervention has no effect on recovery or that delayed rehabilitation may be more beneficial [39,40]. More concerning, early rehabilitation may be detrimental in certain circumstances [41-43]. Clinical trials on acute physical intervention in SCI to date initiate rehabilitation between 2 weeks and 6 months postinjury, bypassing what may be a critical window of recovery very early after SCI [44,45]. To improve understanding of the risks and benefits of very early physical interventions for SCI, we reviewed the preclinical literature on physical interventions delivered within the first 8 days postinjury (Fig. 1).
SUMMARY OF INJURY MODELS AND TRAINING PARAMETERS
To date, the preclinical literature on acute rehabilitation includes studies on rats (n= 46) and mice (n= 6). The most commonly used SCI model is contusion (n= 29), followed by transection (n= 13), and then hemisection (n= 10). Studies using the contusion injury model most often included a moderate impact at mid- to low-thoracic levels and treadmill training as the primary intervention. Of the studies that used transection or hemisection, the majority chose mid- to low-thoracic injuries with bicycle training as the primary intervention. Additional training paradigms included environmental enrichment, ladder stepping, reach training, swim training, exercise ball training, and forced or voluntary wheel running.
Substantial variability exists regarding training dose. Most studies implemented rehabilitation 5 to 7 days per week. Frequency ranged from 1 to 3 bouts of training per day. Duration varied based on intervention type. Treadmill training was delivered between 10 and 40 minutes per day. Bicycle training was most frequently prescribed in two, 30-minute bouts with a 10-minute break between bouts. Other dosages of bicycle training included 10 or 15 minutes per day. Swim training occurred in either 2 bouts of 3 minutes or 4 bouts of 6 minutes per day. Wheel running was prescribed either forced for 20 minutes per day or voluntary, ad libitum access. Reach training typically occurred for 10 minutes per day. For the purpose of this review, we consider voluntary interventions or those performed at a self-selected pace, such as ad libitum wheel running, ball training, reach training, etc. to be “low intensity,” while forced interventions where animals are allowed to rest only at prescribed intervals, like treadmill, cycling, and swim training, are considered “high intensity.”
EFFECTS OF EARLY REHABILITATION ON SCI PATHOPHYSIOLOGY
1. Lesion Pathology, Cell Death, and Apoptosis
To promote functional recovery after SCI, interventions must either limit tissue damage, enhance plasticity of spared or damaged axons, or improve the ability of neuroplastic axons to find appropriate targets. Given that early rehabilitation can alter the spinal microenvironment remote from the injury [46], it is feasible that rehabilitation can modify secondary injury cascades to influence tissue damage at the injury site. Although most studies show little effect of early rehabilitation on lesion parameters, such as tissue damage or white matter sparing [47-54], at least 2 studies suggest potential benefit [36,55]. Indeed, exercise ball training initiated 1 day postinjury preserved tissue caudal to the injury [56], and both cycle training and treadmill locomotion reduced lesion volume when initiated 8 days postinjury [57]. Early rehabilitation may also have favorable effects on lesion pathology when combined with neuroprotective agents, such as polypyrole/iodine mesoparticles [58].
Rehabilitation also has the potential to improve cell survival via caspase inhibition and neuroprotection [59]. Acute passive cycling reduced injury-induced caspase-7 and caspase-9, but not caspase-3 mRNA in the lumbar cord of rats with SCI [59,60]. Surprisingly, caspase mRNA was unaffected by either SCI or acute rehabilitation in the lumbar intermediate laminae and motor neurons [60]. At the protein level, acute cycling reduced both 32-and 40-kD isoforms of caspase-7, the only lumbar caspase increased by injury [59,60]. In large dorsal root ganglia (DRG) neurons, acute rehabilitation decreased injury-induced increases in caspase-3, -7, and -9. When continued long-term after SCI, cycling decreased injury-induced caspase-3 mRNA in the whole cord but increased caspase-3 in motor neurons and both caspase-3 and -9 in large DRG neurons [60]. Given the role of caspases in synaptic plasticity [61], it is possible that late caspase increases serve a nonapoptotic role in response to continued rehabilitation; however, this effect should be further investigated. Overall, it appears that in most instances acute rehabilitation, especially over the short term, decreases caspase expression and may enhance neuroprotective effects during SCI recovery.
Cycling may also selectively influence microRNAs (miRs) involved in apoptosis. For example, miR21, miR16a, and miR217 all decrease caspase expression via inhibition of pro-apoptotic phosphatase and tensin homolog, thus reducing apoptosis [62]. Passive cycling increased miR21 expression in the lumbar cord of rats with SCI but had no effect on miR16a or miR217 following 10 days of training [62]. By 31-day postinjury, no differences were observed for any miRs studied regardless of training suggesting a transient effect early after SCI [62]. Conversely, both early and late cycling reduced the injury-dependent increase of miR199a-3p expression, thereby blocking inhibition of the prosurvival mechanistic (or mammalian) target of rapamycin (mTOR) kinase important for cell survival and proliferation [62].
The effects of rehabilitation on apoptotic pathways suggests that passive cycling is a viable intervention for mitigating aspects of SCI-induced apoptosis early after SCI, despite lesion size and tissue sparing being largely unaffected. Although not directly investigated, rehabilitation effects on mTOR may also reduce necroptosis after SCI [62,63]. Additional research is needed to investigate the role of other rehabilitation interventions on mediators of apoptosis and necroptosis following injury.
2. Plasticity, Sprouting, and Regeneration
Physical interventions rely on repetitive activation of the nervous system to produce effects; therefore, these interventions may have more efficacy for promoting and guiding plasticity than preserving or limiting damage. Several studies investigated early rehabilitation effects on corticospinal tract (CST) sprouting and regeneration given its role in skilled motor functions, but results are, again, mixed [64]. In mice, running wheel exercise specifically enhanced the number of CST collaterals in the thoracic lesion site early after SCI [49]. In the cervical cord, early rehabilitation-induced CST sprouting increases were maintained through chronic time points [49]. In rats, overground locomotor training beginning at 7 days postinjury and using a robotic weightsupport system increased CST sprouting and cortico-reticulospinal bridge formation when combined with an electrochemical prosthesis and treadmill training [65,66]. Two studies assessing CST axon sprouting and length after early reach training also show conflicting results [50,53]. Despite both using rats, similar injury models, and interventions, one study showed significant improvement in CST axon length rostral to the lesion [50] while the other showed no effect of rehabilitation [53]. Immunohistochemistry from another study revealed that following SCI, treadmill trained rats had greater axonal arborization and fiber elongation than sedentary animals [64]. Similar results were found where assisted stepping increased fiber density but not elongation in the CST of rats with SCI [67].
While CST axons are important for skilled movement, serotonergic axons of the raphespinal tract are essential to locomotor function due to their influence on lumbar central pattern generator networks [49]. To date, only one study has investigated serotonergic axon plasticity after early rehabilitation in mice with SCI. In this study, voluntary wheel running did not affect lumbar serotonergic fiber density, but early intervention increased serotonergic contacts on motor neurons in the lumbar cord [49]. By the experiment’s conclusion, both trained and SCI+ sedentary mice exhibited similar lumbar serotonergic reorganization and recovery of rhythmic stepping [49]. While the authors suggest rhythmic stepping can be gained without rehabilitation in incomplete SCI, it is unclear whether the early training-dependent serotonergic fiber reorganization can be exploited by continued rehabilitation to enhance recovery.
Rehabilitation can also modify plasticity of sensory afferents. Indeed, 3 studies histologically quantified calcitonin gene-related peptide (CGRP)-positive fibers in rats due to their role in pain transmission [68-70]. Long-term reduction in CGRP-positive fiber area and length occurred following treadmill training in one study [68] while another study showed no overall effect of training on CRGP fiber density [69]. Passive hindlimb cycling also appears to play a role in reducing CGRP-positive fiber density compared to untrained control rats [70].
Specific acute post-SCI rehabilitation effects on motor neurons remain understudied [71]. While dendritic withdrawal occurred in rats following SCI, acute treadmill training modified these effects by either preventing withdrawal or enhancing extension [71]. Effects occurred primarily in neurons controlling the soleus and to a lesser degree the tibialis anterior muscle [71]. There was no training effect on motor neuron soma size [71].
Perineuronal nets (PNN) are embedded in the extracellular matrix surrounding neurons and facilitate plasticity and maturity of synaptic circuitry [72]. Density of PNNs is greatest in the lumbar region and is both location- and activity-dependent [73]. Both wheel running and treadmill training reversed SCI-induced decreases in lumbar PNNs, but wheel running was most effective in rats [73]. Above the injury, rehabilitation reduced PNNs in gracilis and cuneate nuclei compared to SCI+sedentary mice [73]. Interestingly, treadmill training beginning at 7 days postinjury in rats did not alter PNNs when combined with epidural and pharmacologic stimulation [74]. These findings suggest that rehabilitation differentially modulates PNN expression above and below the injury site to potentially allow greater plasticity in descending systems rostral to the injury while stabilizing lumbar networks susceptible to maladaptive changes and hyperexcitability.
At the synaptic level, treadmill training significantly increased synaptophysin, a synaptic vesicle glycoprotein used to quantify synapses, compared to SCI+sedentary rats [71]. Another study in mice found similar results where treadmill training prevents loss or restores synaptic numbers distal to the lesion [75]. The same effect occurred with postsynaptic density protein 95 [75].
In summary, several preclinical studies demonstrate widespread positive changes in neuron/fiber density and axon growth with rehabilitation. Importantly, no negative effects occurred for structural or neuroanatomical parameters following rehabilitation. These findings support rehabilitation in the first week postinjury as a viable approach to enhance structural plasticity.
3. Neurotransmitter Expression
Neurotransmitters are essential to efficient and effective neural network function. The major inhibitory neurotransmitter, gamma-amminobutyric acid (GABA), regulates synaptic excitability and is also affected by rehabilitation [76]. Magnetic stimulation, treadmill training, and combined treatment significantly increased the number of GABAb-positive cells compared to SCI+sedentary rats [76]. Histologic evidence also suggests that bike and treadmill training may upregulate GABAb and sprouting of dopamine beta-hydroxylase fibers in the ventral horn compared to SCI+sedentary rats [57].
The inhibitory action of GABA is dependent on the intracellular chloride concentration which is heavily influenced by both potassium-chloride cotransporter 2 (KCC2) and sodium-potassium-chloride cotransporter-1 (NKCC1) [77]. Cycling mitigates SCI-induced decreases in lumbar KCC2 following spinal transection in rats [77]. Enriched environment and wheel running increased KCC2 expression compared to SCI+sedentary mice while treadmill training did not [73]. Other sources suggest positive effects of treadmill training where trained rats showed increases in pKCC2/KCC2 ratio and a higher ratio of pKCC2/KCC2 compared to SCI+sedentary rats at all time points [68]. Moderate positive correlations occurred between pKCC2 levels and spasticity, tactile allodynia, and thermal hyperalgesia, and a weak positive correlation occurred between pKCC2 levels and open field locomotion [68]. Cycling also mitigated SCI-induced NKCC1 increases in the rat lumbar cord and maintained KCC2/NKCC1 ratio in the lumbar cord at baseline levels, while SCI+sedentary rats show decreases in this ratio [77].
Additional enzymes that affect GABA neurotransmission are glutamic acid decarboxylase (GAD)-65 and -67 [76]. Spinal cord magnetic stimulation (TMSsc) and treadmill training both significantly increased GAD-67 expression in rats with SCI, but combined TMSsc and treadmill showed the greatest increase compared to individual treatment conditions [76]. Similar results occurred in another study where treadmill trained rats had significantly higher GAD-65 and GAD-67 expression compared to all other treatment groups and did not differ from the uninjured group [78].
Cycling, treadmill training, and treadmill plus magnetic stimulation all enhance neurotransmitter expression following SCI. Due to the necessary role that neurotransmitters play in regulating the efficiency and effectiveness of neural function, implementing rehabilitation during the acute phase of SCI recovery is an appropriate method for enhancing neural communication and facilitating recovery postinjury.
4. Intracellular Signaling Pathways
Rehabilitation may also affect functional outcomes through modulation of intracellular signaling pathways. For example, the phosphorylated form of Erk1/2 kinase, p-Erk1/2, influences neurite outgrowth, survival, and differentiation in both the central and peripheral nervous system and is increased by treadmill training in healthy animals [64]. Recent research also suggests that p-Erk1/2 may enhance regenerative responses in damaged CST axons following treadmill rehabilitation in rats [64]. Indeed, 1 or 2 weeks of treadmill training increased p-Erk1/2 levels significantly beyond injury-induced levels at the injury site and in the motor cortex compared to sedentary rats [64].
The mTOR pathway influences intracellular signaling and regulates protein synthesis, cell growth, and proliferation [62]. Similar to p-Erk1/2, mTOR expression following SCI is affected by rehabilitation [62]. Cycling significantly increased lumbar mTOR levels in rats following 10- and 31-day bouts of rehabilitation [62]. Expression of upstream mTOR positive mediators, including transforming growth factor alpha and phosphoinositide 3 kinase in the lumbar cord was significantly increased following cycling [62]. Additionally, positive downstream effectors, such as S6K1 and S6, were not affected by SCI but still increased following rehabilitation intervention [62], while downstream inhibitory effector, eukaryotic initiation factor 4E but not eukaryotic translation initiation factor 4E binding protein 1 was decreased by shortterm rehabilitation [62].
In summary, there is clear evidence of positive intracellular signaling changes in response to rehabilitation early after SCI. Rehabilitation during acute recovery upregulates expression of cell signaling molecules that promote neurite growth, survival, and differentiation as well as protein synthesis, cell growth, and cell proliferation. Each of these molecular changes has the potential to enhance functional outcomes and should be targeted by prescribing treadmill or bicycle training during acute recovery.
5. Neurotrophic Factors, Growth Factors, and Their Receptors
Neurotrophic and growth factors enhance survival, maintenance, and synaptic strength and regulate processing of sensory and motor stimuli [48,79]. Rehabilitation is a noninvasive means of increasing these factors, thereby mitigating neuropathic pain and other potential deficits [48].
Brain-derived neurotrophic factor (BDNF) is a multifunctional protein with wide-ranging effects, including acting as a chemo-attractant for regenerating axons [60]. Running wheel exercise increased BDNF in the ipsilateral hemicord at all time points compared to uninjured and SCI+sedentary rats [80]. Passive cycling also increased BDNF in the spinal cord, motor neurons, and intermediate grey matter, but not the DRGs of rats that received short- and long-term rehabilitation compared to uninjured and SCI+sedentary rats [60]. Treadmill, swimming, and standing all increased BDNF levels compared to SCI+sedentary rats; however, only treadmill training maintained soleus BDNF levels comparable to uninjured animals. All other groups showed levels below baseline [51]. Wheel training produced mixed results for BDNF protein expression where some groups’ expression surpassed controls while others showed no significant results [80,81]. Treadmill training enhanced BDNF protein levels [68,78,79]; however, combining treadmill with magnetic stimulation showed the greatest increases [76]. In contrast, cycle training increased BDNF protein expression in several studies, but only some studies showed long-term benefit [60,77,79,82,83].
Tropomyosin receptor kinase B (TrkB), the BDNF receptor, regulates its ability to function [68]. In rats, 10 days of passive cycling increased TrkB mRNA expression in the whole cord, specifically in motor neurons. These effects were not observed after 31 days of training [60]. There was no training effect in the intermediate grey matter or large DRG neurons [60]. Treadmill and passive cycle training significantly increased TrkB protein expression in rats with SCI in several additional studies as well [60,68,77,78], while bike training had no effect [82]. Interestingly, early rehabilitation did not affect TrkB signaling effector phospholipase Cgamma or the TrkB docking site, Shc [68]. Wheel running in rats with SCI normalized injury-induced decreases in cyclic AMP response element binding protein (CREB) mRNA and protein, which mediates downstream expression of TrkB, although no effects on phosphorylated CREB expression were found at the conclusion of the study [80]. Treadmill trained rats had higher levels of CREB in the dorsal horn compared to SCI+sedentary and TrkB-blocked rats [78].
Synapsin I is downstream of BDNF and influences neurotransmitter release, axonal elongation, and synaptic maintenance [80]. Voluntary wheel running increased synapsin I mRNA compared to SCI+sedentary rats [80]. Swimming and standing showed greater benefit in the lumbar cord than treadmill training [51]. Standing trained rats also showed significant improvement versus SCI+sedentary rats while swimming showed nonsignificant improvement [51]. In the soleus muscle, synapsin I was also significantly increased by standing training compared to SCI+sedentary, treadmill, and swimming groups [51].
Glial cell-derived neurotrophic factor (GDNF) modifies neural circuitry by enhancing strength and promoting synapse development [60]. Cycle-trained rats showed increased GDNF mRNA in the whole spinal cord, motor neurons, and intermediate grey matter compared to uninjured and SCI+sedentary rats; however, no differences occurred in large DRG neurons for any group [60]. Protein levels also significantly increased following wheel and step training compared to SCI+sedentary groups [48,79]. Short- and long-term passive cycling increased whole cord RET expression, a cell signaling receptor for GDNF [60]. Passive cycling also increased another GDNF receptor, GFRα1, in large DRG neurons but not in the whole cord or motor neurons following SCI [60].
Neurotrophin-3 (NT-3) is another factor that mediates synaptic transmission and regeneration [80]. Associated with sensory neuron survival and function, NT-3 deficits can lead to sensory neuron loss and subsequent gait dysfunction [80]. Voluntary wheel running in rats with SCI restored injury-induced NT-3 mRNA levels after hemisection [80]. Further, passive cycling for 10 days increased NT-3 mRNA in the whole spinal cord, motor neurons, and intermediate grey matter while a 31-day regimen showed no significant effects [60]. No rehabilitation effects were seen in large DRG neurons with either paradigm [60]. Treadmill and swimming rehabilitation significantly increased NT-3 mRNA in the lumbar cord compared to control, but only treadmill training showed significant increases compared to sedentary rats [51]. Soleus NT-3 mRNA was significantly increased following treadmill, swimming, and standing training compared to SCI+sedentary rats; however, treadmill training was the only approach that significantly increased levels beyond uninjured values [51]. Protein levels were increased by passive cycling and stepping but not wheel running exercise [60,79,80].
Tropomyosin receptor kinase C (TrkC), the receptor for NT-3, showed increased protein expression in the whole cord of rats who received passive cycling intervention compared to SCI+sedentary rats after SCI [60]. No differences were found for TrkC mRNA expression in the whole cord, motor neurons, or large DRG neurons following injury or rehabilitation [60].
Similar to those above, neurotrophin-4 (NT-4) functions as a chemo-attractant for regenerating axons while strengthening and enhancing synapse formation [60]. Step training significantly increased NT-4 mRNA and protein in rats with spinal cord transection [79]. Passive cycling also significantly increased NT-4 protein expression in transected rats [60]. Whole cord expression of NT-4 mRNA was not affected by passive cycling; however, NT-4/5 mRNA expression was increased in motor neurons and intermediate grey matter but not large DRG neurons following transection [60].
Growth associated protein 43 (GAP-43) is expressed in growing axon terminals and influences growth, neurotransmitter release, learning, and memory [80]. Following SCI, GAP-43 expression is often decreased, but rehabilitation may reverse these effects [80]. Indeed, training-induced normalization occurred following wheel running in rats with SCI [80]. Reach training significantly increased GAP-43 expression contralesional to the preferred paw compared to injured, untrained rats and ipsilesional to the preferred paw [50]. Bike training did not affect GAP-43 expression in rats [82]; however histologic evidence suggests that passive cycling increased the percentage of GAP-43 positive processes in the lesion area [83].
In summary, various training paradigms including wheel, step, treadmill, cycling, and swimming training have the potential to induce molecular changes in spinal trophic and growth factor expression following SCI. Differential results suggest that early rehabilitation largely upregulates neurotrophic and growth factor expression which in turn has the potential to enhance synapse formation, survival, and strength. Factor expression in DRG neurons is largely unchanged by rehabilitation, potentially due to inherently high intrinsic capacity for growth and repair [84]. Utilizing rehabilitation as a means of enhancing neuroplasticity through synaptic changes may be another viable target for promoting functional recovery in this population.
6. Inflammation
Inflammation can both positively and negatively influence recovery following injury [85-89]. As a crude marker for microglial and macrophage activation state, ionized calcium-binding adapter molecule 1 (IBA1) is often used to assess inflammatory response following SCI [73]. Enriched environment and voluntary wheel running initiated 1 week postinjury significantly increased IBA1 expression at L5 compared to uninjured, while treadmill trained and SCI+sedentary groups showed no difference from control in mice with thoracic contusion [73]. No difference in lumbar IBA1 expression occurred for any group [73]. The same study showed that wheel running and treadmill training significantly decreased microglial/macrophage isolectin B4 (IB4) expression while enriched environment and sedentary mice showed no difference from control [73]. Wheel running initiated at 5 days postinjury that continued for 4 weeks did not influence C7–8 dorsal horn IBA1 expression in rats with cervical contusion injuries [47]. Treadmill training beginning 2 days postinjury shifted the lumbar microenvironment toward a more proinflammatory state by increasing mRNA of inflammatory and trafficking genes while decreasing proreparative genes of infiltrating macrophages in the mouse lumbar cord. This occured without influencing the total number of infiltrating cells in the spinal cord or DRG [46,47].
Similar to IBA1, glial fibrillary acidic protein (GFAP) is a classic marker used to assess astrocyte reactivity [73]. In rats, passive cycling at 5 days postinjury increased GFAP levels after both 10 and 31 days [60]. Enriched environment was also shown to enhance GFAP levels at L6 compared to uninjured, SCI+sedentary, and wheel trained mice in a separate study; however, L5 GFAP levels were significantly higher in all treatment groups compared to control [73].
Currently, it appears that intense rehabilitation initiated too early can negatively impact inflammation during SCI recovery. Further research is needed to conclude whether changes in microglia, macrophage, or astrocyte responses confer the negative impacts of hyperacute treadmill training or if other cell types are involved. These results would help better understand the risks and rewards of implementing rehabilitation during the hyperacute phase of recovery.
7. Muscle Tissue
Muscle is essential for functional capacity and an important source of metabolic support that undergoes tremendous change after SCI [90]. Early treadmill training increased cross-sectional area in the soleus muscle compared to untrained animals [75,91,92]. Although not statistically significant, 5 days of treadmill training showed a trend toward attenuation of mixed I+IIa fibers and prevented the appearance of type IIa+IIx fibers compared to SCI+sedentary rats [91]. Fatigue analysis showed significantly lower fatigue levels in the treadmill trained rats compared to the SCI+sedentary group [91]. Despite changes in cross-sectional area and fatigue, no significant difference was found for twitch force or peak tension in the soleus muscle between uninjured, SCI+treadmill, or SCI+sedentary rats [91].
Muscle growth and regeneration are affected by myogenic regulatory factors influenced by rehabilitation [92]. Although protein levels remained unchanged, myogenic differentiation 1 mRNA was significantly increased in SCI+sedentary rats at 14 days postinjury while trained rats showed significant increases in expression at 8 and 14 days postinjury compared to uninjured [92]. Treadmill training significantly increased myogenin mRNA and protein levels compared to uninjured rats [92] and those with SCI alone [93]. Expression of myogenic factor 5 mRNA was not influenced by treadmill training in rats with contusion SCI; however, protein expression was significantly increased [92]. Embryonic myosin demonstrates a muscle-specific response to early rehabilitation after SCI. In the soleus, treadmill training significantly increased embryonic myosin compared to SCI+sedentary and uninjured treadmill trained rats [93]. In contrast, embryonic myosin in the tibialis anterior of SCI rats was significantly greater compared to the tibialis anterior of all other groups [93].
Insulin-like growth factor 1 (IGF1) also alters muscle mass and repair [92]. Overexpression of IGF1 promotes hypertrophy, and under expression can cause muscle fiber abnormalities [92]. At 14 days postinjury, SCI+sedentary rats had significantly higher IGF1 expression compared to control rats, while treadmill trained SCI rats showed significantly higher expression compared to uninjured and SCI+sedentary rats [92]. Similar results were found for IGF1 peptide [92]. Additionally, higher IGF1 peptide concentration was found in the rat soleus muscle compared to tibialis anterior for uninjured, uninjured+treadmill, and SCI+treadmill groups [93]. Treadmill trained rats had significantly higher IGF1 peptide in the soleus than all other groups [93]. Receptor expression was also increased in trained rats compared to uninjured and SCI+sedentary rats following 14 days of training [92]. By 8 and 14 days postinjury, treadmill trained rats showed a 10-fold increase in IGF1B mRNA compared to uninjured and SCI+sedentary rats while SCI+sedentary rats showed a significant increase in mechano-growth factor mRNA, an IGF isoform, compared to uninjured only at 14 days postinjury [92].
A family of IGF binding proteins (IGFBP) influence IGF1 activity [92]. This family includes 7 different binding proteins, but only IGFBP4 and IGFBP5 affect muscle fiber changes [92]. Treadmill trained rats showed higher levels of IGFBP4 after 8 and 14 days compared to uninjured and SCI+sedentary rats; however, IGFBP5 was significantly higher only after 14 days of treadmill training [92].
Rehabilitation enhances muscular adaptation and strength in rodents with SCI. Specifically, data supports the use of treadmill training within the first 8 days postinjury to enhance muscle regulatory factors, increase cross-sectional area, and decrease fatigue. Using acute treadmill training to target muscle fiber adaptation can facilitate functional recovery by mitigating SCI-induced strength deficits.
EFFECTS OF EARLY REHABILITATION ON CLINICAL OUTCOMES
1. Gross Motor Function
Earlier admittance to acute care facilities and earlier time to rehabilitation following SCI has the potential to improve patients’ functional ability for tasks including, but not limited to, standing, bathing, grooming, and balance but does not affect scores on the Walking Index for Spinal Cord Injury [38,94,95]. Similar results were found in rats where early rehabilitation involving reach, wheel, and treadmill training showed improvements in motor function based on Forelimb Movement Score, Forelimb Locomotor Score, and Motor Function Score [53,64,81]. Importantly, task-specific reach training resulted in significant reaching improvement but significantly worsened performance on ladder tasks in the same group of rats, highlighting the potential for interference with skill learning [50].
Regarding hindlimb motor function, treadmill training yields an overwhelmingly positive response on open field locomotor scores at early and late time points after injury [36,52,57,58,64,68,69,71,73,75,91]. Treadmill trained rats also showed significant reduction in muscle coactivation in ankle flexor and extensors and spasticity compared to SCI+sedentary rats [68]. Only 2 studies showed no significant difference in open field locomotor performance after early treadmill training [41,55]. In fact, Hansen et al. [41] observed a significant positive effect of early treadmill training in mice, but only when inflammation was controlled. Use of exercise balls for activity following SCI showed significant improvement in open field locomotor scores compared to delayed training in rats; however, performance in all groups plateau roughly 2 weeks postinjury [56]. Passive hindlimb cycling, swimming, wheel running, and enriched housing had no effect on open field locomotor scores in mice or rats following SCI [36,81,96,97].
Stepping analysis revealed that rats trained on a horizontal ladder task immediately after injury had significantly greater stepping ability after 1 week of training compared to those who received delayed training [54]. Additionally, a strong positive correlation was found between damage in the thoracic dorsal and dorsolateral funiculi at T8 and initial error rate for stepping in early and delayed training groups [54]. Ladder task performance significantly improved when flat running wheels were used for mice instead of traditional, runged wheels and for rats with lateral funiculus and dorsal funiculus lesions but not entire dorsolateral quadrant lesions [36,53]. Early and late voluntary wheel training elicited significant improvement in Rotarod performance and decreased the number of foot falls on a ladder task compared to baseline in mice with dorsal hemisection [49]. Wheel trained mice also covered significantly greater distances on a running wheel assessment compared to treadmill trained mice [73]. Hindlimb or locomotor-type rehabilitation did not affect grip strength in rats with SCI [81].
The current body of literature suggests that the effects of early rehabilitation on gross motor recovery following SCI are largely positive with only one study reporting unwanted outcomes associated with rehabilitation. Negative outcomes in this study may be attributed to the use of high-intensity rehabilitation initiated 2 days postinjury, at which point the injury may be too dynamic for subjects to tolerate this level of exertion unless inflammation is controlled [41]. In contrast, treadmill, wheel, and cycle training initiated later during the acute phase following SCI are highly effective and should be used to promote gross motor recovery.
2. Fine Motor Performance and Kinematic Analysis of Function
Evidence from footprint analysis of treadmill trained mice showed improvement in hindlimb alternation and stride length; however, smearing of hindlimb pawprints at push-off and absent heel contact were also noted [75]. In contrast, enriched housing, wheel running, and treadmill training had no effect on maximal foot contact or footprint score following SCI in rats [55,97]. Kinematic analysis of mouse joint angles and limb movement showed that treadmill training improved hip and ankle joint movement with decreased variability in foot posture [75]. Treadmill training also decreased the angle of rotation in the feet following contusion injury in rats [57]. Combined treadmill and TMSsc in rats mitigated SCI-induced increases in wrist but not ankle lift off angle compared to control and TMSsc only groups [76]. Cycling and treadmill training in rats showed improvement in base of support, while running wheel and enriched housing had no effect [57,97]. Wheel running, enriched housing, and treadmill training also had no effect on thoracolumbar height, maximal leg extension, or yield during stance following SCI in rats [55,97]. Reach training showed improvement on reaching tasks without changing reaching style [50,53].
Treadmill training provides the most benefit regarding changes in fine motor performance and kinematics in animals with acute SCI, while enriched housing and wheel running are largely ineffective. To promote recovery of fine motor skills following SCI, treadmill training or task-specific reach training should be the first-choice intervention; however, careful consideration of dosage timing and intensity is still needed to maintain safety and maximize benefit.
3. Sensory Function
Hutchinson et al. [51] found that treadmill, swimming, and standing training all significantly increased time to peak flexion withdrawal following noxious pinch stimulus to the rat hind paw; however, only treadmill trained rats showed significant improvement in mechanical sensitivity following contusion injury [51]. Treadmill training also improved mechanosensory thresholds in rats receiving more severe contusion injury in 2 of 3 models that investigated these effects [68,71,78]. Rats trained with exercise balls immediately following injury showed significantly higher tactile withdrawal thresholds compared to those that received delayed training [56]. Importantly, delayed training did not reverse allodynia once it had developed [56]. Five weeks of wheel running initiated at 5 days postinjury decreased prevalence of at-level allodynia from 42% to 6% in rats with SCI [48]. Moreover, rats that received rehabilitation throughout the remainder of the study did not develop allodynia in the ipsilateral or contralateral forepaw [48].
Early treadmill training effects on thermal withdrawal thresholds after SCI show mixed results. Some studies in rats show significant improvement in withdrawal threshold in response to thermal stimuli [68,78], while others show no effect of rehabilitation [48,56,71]. Although there was variation in training paradigm and injury models, both studies that found positive effects on thermal withdrawal threshold utilized body weight-supported treadmill training [68,78]. Running wheel exercise also restored thermal withdrawal latencies to baseline levels in mice, while enriched housing had no significant effect [73].
Various paradigms including treadmill, swim, stand, wheel, and exercise ball training enhance at least one aspect of sensory function following SCI. Despite this, treadmill and wheel training are the most effective for promoting recovery of sensory processing and should be prioritized when choosing interventions. Effects of rehabilitation on thermal processing are not as clear, but treadmill and wheel training show benefit in several studies and should be prioritized over enriched environment.
4. Reflex Function
Following SCI, supraspinal control is diminished while afferent feedback becomes augmented [79]. These changes lead to decreased spinal reflex modulation and can cause hyperreflexia, spasticity, and inappropriate timing of movement [79]. To investigate reflex changes following SCI, numerous studies have assessed H-reflex modulation and Hmax:Mmax ratio changes, a relative indicator of motoneuronal excitability [79].
Frequency-dependent H-reflex depression was enhanced in bike and step-trained rats after 30 days of training [79]. No differences were found for Hmax:Mmax ratio in response to increasing stimulus intensity beyond the motor threshold; however, slope of the recruitment curve was significantly higher in step-trained rats compared to bike-trained and untrained groups [79]. A subsequent study by the same group showed that spinal excitability in SCI+sedentary rats had significantly increased Hmax:Mmax ratios in the first 2 weeks following injury that plateaued around 28 days postinjury [77]. Implementing early cycle training restored spinal excitability at 14 and 28 days postinjury by increasing frequency-dependent modulation of the H-reflex [77]. Rehabilitation also significantly reduced Hmax:Mmax ratio by 28 days postinjury [77]. Additional research in rats also supports the idea that frequency-dependent depression is improved with bicycle and treadmill training [57,73].
Wheel training in mice showed significant decreases in Hmax: Mmax ratio over time while other treatment groups did not [73]. Additionally, the percentage of rate-dependent depression as a function of frequency showed that wheel trained animals did not differ significantly from control animals while sedentary, enriched housing, and treadmill trained groups all showed significantly higher values when the experiment concluded [73].
The current body of research suggests that various training paradigms, including treadmill, bike, and wheel training, mitigate SCI-induced decreases in H-reflex modulation and simultaneously reverse increases in the Hmax:Mmax ratio following injury. These findings show only positive effects of early rehabilitation on spinal excitability and reflex recovery after injury and should be utilized readily in future studies.
5. Autonomic Function
The effects of SCI extend beyond somatic functions to include numerous autonomic and systemic changes [98-100]. Early rehabilitation has the potential to enhance protective effects on cardiac indices at early time points following SCI in rats, but only if rehabilitation is continued over time [101]. Rehabilitation initiated at later time points can still elicit positive effects, but subsequently increases time to recovery [101]. In rats, resting heart rate and heart rate responses to colorectal distension-induced autonomic dysreflexia are largely unaffected by treadmill training and passive hindlimb cycling following SCI while changes in systolic blood pressure (SBP), mean arterial pressure (MAP), and the severity and frequency of autonomic dysreflexia show varied results between studies [69,70,101]. Indeed, one study found that treadmill trained animals had increased MAP during training and significantly decreased MAP following training [69] while another showed that MAP was unaffected by passive cycling initiated early or late after injury [101]. Treadmill trained rats also had larger MAP responses during periods of autonomic dysreflexia compared to SCI+sedentary rats [69]. Delayed passive hindlimb cycling increased the pressor response to colorectal distention at 5 but not 9 weeks postinjury and decreased the incidence of autonomic dysreflexia by the study’s conclusion [101]. Additionally, West et al. [70] found that passive hindlimb cycling reduced the SBP response to colorectal distention while Popok et al. [101] showed no difference between groups at any time point for pressor response during autonomic dysreflexia, or maximal SBP reached. Early initiated treadmill training has little to no effect on vascular conductance following rat spinal cord transection [69] while swim training initiated at 3 days postinjury increased microvascular dysfunction [102].
Changes to the urinary system in response to early intervention show that locked and unlocked flat surface running wheel groups were not different in the number of days to micturition, while the rung wheel group showed longer time to micturition compared to uninjured and SCI+sedentary mice [36]. A trend between higher open field locomotor scores and earlier micturition was described, but significance and correlational values were not included [36].
Data show that treadmill training initiated within the first 3 days after injury negatively impacts cardiovascular function while bicycle training initiated later in the acute phase of recovery has only positive effects. Continued research is needed to determine effects of bicycle training within the first 3 to 4 days after injury to ascertain whether cycle training is genuinely more beneficial than treadmill training during this period. Treadmill training should be avoided within the first 4 days of injury to protect cardiovascular function; however, other high-intensity interventions such as bicycle and swim training should also be utilized with caution.
EFFECT OF EARLY TRAINING PARAMETERS ON OUTCOMES
Four of the 9 studies that initiated rehabilitation within the first 3 days found negative effects regarding autonomic dysreflexia, inflammation, microvascular dysfunction, and longterm locomotor outcomes [41,46,69,102]. Additionally, 3 of the studies that showed negative effects used treadmill training in their rehabilitation paradigm [41,46,69]. Two of these studies implemented body weight-supported treadmill training with manual paw placement in two, 10-minute bouts separated by 20-minute rest periods for roughly 1 week following injury in moderately contused mice and showed worsened locomotor function [41,46]. The other used 10 minutes of treadmill training per day, 5 days per week for 6 weeks which worsened autonomic dysreflexia but improved open field locomotion in rats with transection injury [69]. The fourth and final study implemented swim training 3 days after SCI in rats and showed worsened microvascular dysfunction [102].
A single study that implemented treadmill training between 2 and 4 days postinjury showed positive effects on function using three, 10-minute bouts of treadmill training with a 5-minute break in between bouts at 5 days per week for 12 weeks in rats with microballoon compression injuries [52]. Results of this study showed improved open field locomotion at 2 weeks that plateaued at 7 weeks but remained higher compared to sedentary animals [52]. Another study implemented 15 minutes of daily treadmill training beginning 4 days postinjury and showed slightly larger exploratory behavior but no additional benefit on locomotor outcome [55]. Other studies that started rehabilitation in the first 3 days without negative effects used ladder stepping, exercise balls, and voluntary wheel training [49,54,56].
Continuing rehabilitation beyond the first week postinjury may overcome the negative effects of treadmill training within the first 3 days [52,69]. Both the studies of Hansen et al. [41] and Norden et al. [46] implicate enhanced inflammation as a potential cause for negative outcomes from early rehabilitation. Although Multon et al. [52] and Laird et al. [69] did not investigate inflammatory changes, they saw locomotor improvement after 6 and 12 weeks of continuous training, respectively. It is important to note that Laird and colleagues saw significant, nonfunctional gains in locomotor performance which may be related, in part, to the administration of nonsteroidal anti-inflammatory drugs during recovery [69].
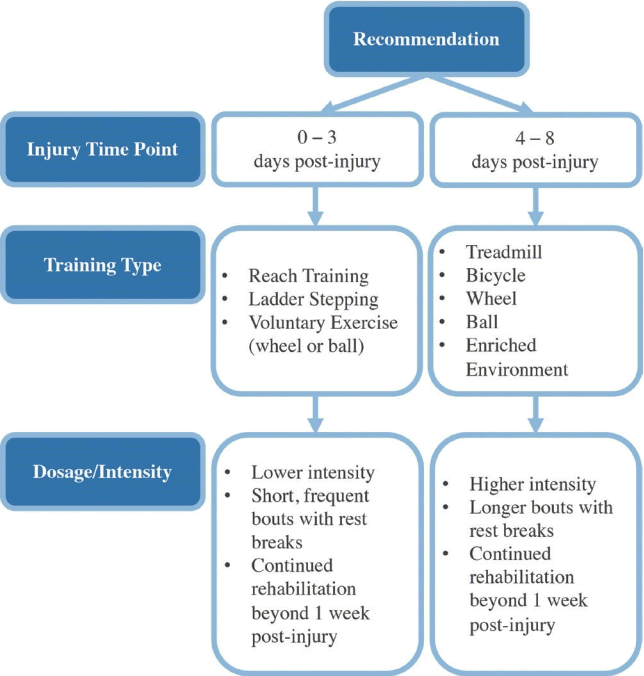
In summary, effects of rehabilitation during the hyperacute phase of SCI are largely dependent on injury and rehabilitation paradigms. It appears that lower-intensity or self-paced rehabilitation, such as reach or ladder training, in shorter, more frequent bouts elicits better outcomes when implemented between 0- and 3-day postinjury. These benefits may be attributed to less inflammation resulting from lower rehabilitation intensity, but additional research is needed to confirm this. Higher-intensity rehabilitation is more beneficial when started after 4 days of recovery and can be dosed in longer bouts with less frequent rest breaks. Despite differences in dosage and intensity, rehabilitation initiated during the hyperacute phase should continue past the 1 week recovery mark to ensure that all short- and longterm effects are recognized (Fig. 2).
CONCLUSION
Treadmill training is the most studied rehabilitation approach in the first week post-SCI and shows the greatest effect on locomotor outcomes. Independent of training parameters, cycle training shows the least risk of adverse effects and is the topic of an ongoing clinical trial [103]. However, no studies implemented cycle training within the first 3 days after injury and is a major gap in the literature. Early physical activity is most beneficial for synaptic plasticity, structural plasticity, neurotransmitter/neurotrophic/growth factor expression; least effective for tissue sparing; and potentially detrimental if too intense too soon under uncontrolled inflammation.
To promote positive outcomes during acute SCI recovery, lowintensity or self-limiting rehabilitation such as reach training or voluntary wheel or ball exercise should be utilized for the first 3 days postinjury. Activity dosage during this period should include short, frequent bouts of rehabilitation with rest breaks, and the duration should extend long-term to maximize benefit. Higher-intensity rehabilitation including treadmill, bike, and enriched environment can be implemented safely from 4 days postinjury onward. Rehabilitation at this point can be dosed in longer bouts with less frequent rest breaks but should also extend beyond the first week of recovery (Fig. 2). It is important to recognize that these data and recommendations are limited by the discrepancy between animal and human biological and recovery timelines. Comparative biological studies are warranted to identify cross-species timelines for effective translation of these findings to humans.
While we have a general idea which interventions and dosages are safe and effective in acute SCI, data in the hyperacute phase are scarce. As such, future research should focus on identifying which interventions are safe and most effective during the hyperacute phase; how to best manipulate the hyperacute window to allow greater intensity intervention; multisystem effects of spinal cord stimulation or bicycle training during the first 3 days postinjury; whether interventions other than passive cycling can reduce post-SCI apoptosis; and the effect of acute rehabilitation on nonmyeloid cell function. Given the tremendous beneficial effects of physical rehabilitation in the acute phase, particular emphasis should be placed on combining emerging therapeutics, particularly those targeting regenerative or inflammatory responses, with acute rehabilitation.
Lastly, substantial variability in injury models, rehabilitation paradigms, and chosen outcomes makes systematic review of the literature, drawing definitive conclusions, and thus making recommendations challenging. Some degree of standardization within the field may speed knowledge development and clinical translation. As such, increased focus on clinically relevant injury models, such as contusion, is warranted. It is also prudent for basic and clinical scientist in SCI research to collaborate in developing, standardizing, and validating a translational battery of measures spanning behavioral, electrophysiological, and imaging modalities to assess outcomes across species. It is our hope that this work and recommendations can be used to guide future systematic reviews of the literature and facilitate these discussions. Investigating these effects along with implications for the areas mentioned previously would greatly benefit our ability to understand and prescribe effective rehabilitation during this delicate stage to enhance long-term recovery.